饥饿胁迫对日本蟳抗氧化酶活性和细胞超微结构的影响
时间:2022-12-08 09:25:02 来源:雅意学习网 本文已影响 人 
丁爱侠
饥饿胁迫对日本蟳抗氧化酶活性和细胞超微结构的影响
丁爱侠
(宁波大学 土木与环境工程学院, 浙江 宁波 315211)


日本蟳()隶属于梭子蟹科、蟳属, 浙江俗称“石蟹”“石奇角”和“石蜞螃”, 在日本、朝鲜、马来西亚等国家广泛分布, 国内渤海、黄海、东海、南海等海域也有分布. 因其营养价值高、肉味鲜美, 深受消费者喜爱, 具有较高的经济价值[1-2]. 然而, 近年来海洋日本蟳产量越来越少, 人工增养殖、人工放流逐渐成为餐桌上日本蟳的主要来源. 因此, 日本蟳基础生物学的研究成为日本蟳增养殖产业化发展以及人工放流规模扩大的重要理论支撑.
自然放流环境、养殖环境以及运输过程中的水生生物很容易受到饥饿的胁迫, 体内免疫系统和细胞结构发生改变, 对各种病原生物的抵抗力降低, 从而导致疾病暴发. 国内外学者先后研究了饥饿胁迫对蓝鳃鱼、克氏原螯虾、罗非鱼、刀鲚、团头鲂、猛虾蛄等水生动物抗氧化酶活性和器官组织结构的影响[3-8]. 但是, 国内外关于环境胁迫对于日本蟳免疫系统和细胞结构、功能影响的研究成果相对较少, 尚未见到饥饿胁迫对日本蟳组织细胞和酶活性影响的相关报道.
本研究选择超氧化物歧化酶(superoxide dismutase, SOD)和过氧化氢酶(catalase, CAT)作为研究指标, 通过试验探讨日本蟳在饥饿胁迫下体内抗氧化酶活性的变化, 分析日本蟳生理调节机制. 同时运用组织学试验方法, 对日本蟳鳃、肌肉、肝胰腺、心脏的细胞超微结构进行拍照, 观察16d饥饿胁迫后细胞结构的变化, 为加强日本蟳健康养殖调控、改进活体运输技术以及丰富日本蟳生态学理论研究提供试验数据支撑.
1.1 试验材料
日本蟳购自宁波象山码头, 共200只, 雌雄各半, 规格整齐((6.8~7.2)cm×(4.6~5.3)cm). 日本蟳购入后放入生态循环养殖系统暂养, 试验所用海水为鄞州育苗场经沉淀和过滤后的清洁海水, 盐度2.0%, 水温10℃, 自然光照, 连续充气. 暂养期间每日2次投喂新鲜贝类, 并仔细观察, 淘汰缺少活力的个体, 暂养7d后开始饥饿胁迫试验.
1.2 试验设计
饥饿胁迫试验养殖条件、养殖方法同暂养, 本试验设2个对照组和2个饥饿组, 每组随机放入30只预先挑选出来的体态正常、健康、活力强的个体, 雌雄各半. 对照组正常投喂新鲜贝类, 饥饿组不投饵. 16d后终止试验.
1.3 试验方法
1.3.1 抗氧化酶活性的测定
对照组和饥饿组每4d取样1次, 每次取2~3只, 迅速置于冰盘内活体解剖, 分别取出鳃、肝胰腺、心脏和肌肉, 用生理盐水冲洗后放入-70℃冰箱备用. 测定时准确称量0.1g组织样品, 加入0.5mL磷酸盐缓冲液(pH=7.0), 放入玻璃匀浆器, 在冰浴条件下匀浆5min, 然后放入冷冻离心机内以14000r·min-1的速度离心25min, 取上清液用于酶活性测定.
SOD活性测定采用氮蓝四唑光化学反应法[9]. 3mL反应液中含50mmol·L-1磷酸缓冲液(pH=7.8) 1.5mL, 130mmol·L-1甲硫氨酸(met)溶液0.3mL, 0.75mmol·L-1氯化硝基四氮唑兰(NBT)溶液0.3mL, 0.1mmol·L-1EDTA-Na2液0.3mL和组织上清液0.1 mL, 最后加0.02mmol·L-1核黄素0.3mL. 置于4000lx日光下进行光化学反应, 反应温度20℃, 30min后用黑暗终止反应, 并立即使用DR5000分光光度计(哈希, 美国)在560nm的波长下测吸光度. 一个SOD活力单位定义为能引起反应初速度(指不加酶时)半抑制时的酶用量. CAT活性测定采用紫外吸收法. 3mL反应液中含0.2mol·L-1磷酸缓冲液(pH=7.0) 1.5mL, 组织上清液0.2mL和0.1mol·L-1H2O20.3mL. 混合后即于240nm的波长下测吸光度, 然后置于25℃水浴中反应5min, 再置于240nm下测定吸光度. 根据前后吸光度的差值计算CAT活性, 以1min内吸光度减少0.1的酶量为1个酶活单位. 每组样品各设置4个平行.
1.3.2 电镜观察
对解剖获得的鳃、肌肉、肝胰腺和心脏组织迅速用2.5%戊二醛和1%锇酸双固定, 梯度乙醇脱水, Epon812环氧树脂包埋, 以LKB-3型超薄切片机(LKB, 瑞典)切片, 醋酸铀和柠檬酸铅双染色, 用H-700型透射电镜(日立, 日本)观察并摄影.
1.4 数据分析
采用SPSS 17.0统计软件分析酶活性数据, 采用均值Duncan法进行差异性比较, 显著水平设为<0.05.
2.1 饥饿胁迫对日本SOD和CAT活性的影响
试验结果表明两种抗氧化酶在日本蟳体内的分布具有组织差异性. SOD酶的活性大小顺序为: 心脏>鳃>肌肉>肝胰腺, CAT活性为肝胰腺>心脏>鳃>肌肉. 两种酶在饥饿胁迫下表现出不同的应激反应.
饥饿胁迫对日本蟳SOD活性的影响见表1. 鳃组织中SOD活性在饥饿胁迫第4d开始增加, 第8d显著降低, 第12d又快速升高, 并在试验结束前保持一个稳定的高水平. 肝胰腺组织中SOD活性在饥饿胁迫的前8d内较对照组有轻微的下降, 第12d突然升高, 并达到峰值后稳定至试验结束. 心脏和肌肉组织中SOD活性总体受到饥饿胁迫的影响相对较小, 其中心脏组织中SOD活性在饥饿胁迫下呈现比较明显的刺激作用, 肌肉组织中SOD活性在第4d表现出轻微的抑制作用, 从第8d开始到结束一直维持在刺激状态. 总体看来, 饥饿胁迫会让日本蟳机体内SOD活性表现出一定的刺激效应, 并且在第12d达到峰值.

表1 饥饿胁迫对日本SOD活性的影响
注: 数据为平均值±标准偏差, 不同上标字母表示同一组织中存在显著性差异(<0.05). 下表同.
饥饿胁迫对日本蟳CAT活性的影响见表2. 鳃中CAT活性在试验期间的波动较小, 且整体表现出轻微的抑制作用. 肝胰腺中CAT活性对饥饿的反应比较敏感, 试验第4d出现明显的抑制作用, 酶活性只有对照组的40%, 第8d开始快速增加达到峰值, 12d恢复并接近对照组的水平, 第16d酶活性略有回升. 饥饿胁迫第4d肌肉CAT酶活性出现明显的应激反应, 升高到对照组的2倍, 其余时间基本保持在对照组水平. 饥饿对心脏CAT活性影响相对较小, 16d内基本维持在对照组水平.
表2 饥饿胁迫对日本CAT活性的影响
2.2 饥饿胁迫对日本组织超微结构的影响
如图1所示, 对照组日本蟳鳃丝管壁比较厚, 由上皮细胞构成, 外层为角质层; 细胞质中内质网、线粒体都十分丰富; 细胞核规则, 核膜清楚, 异染色质丰富、染色深. 饥饿胁迫组日本蟳鳃丝的管壁轻微变薄; 部分鳃丝出现轻微肿胀; 线粒体部分解体, 但仍有一部分比较完整, 内嵴水肿、解体, 并有空泡化现象出现; 内质网和细胞核都比较完整.
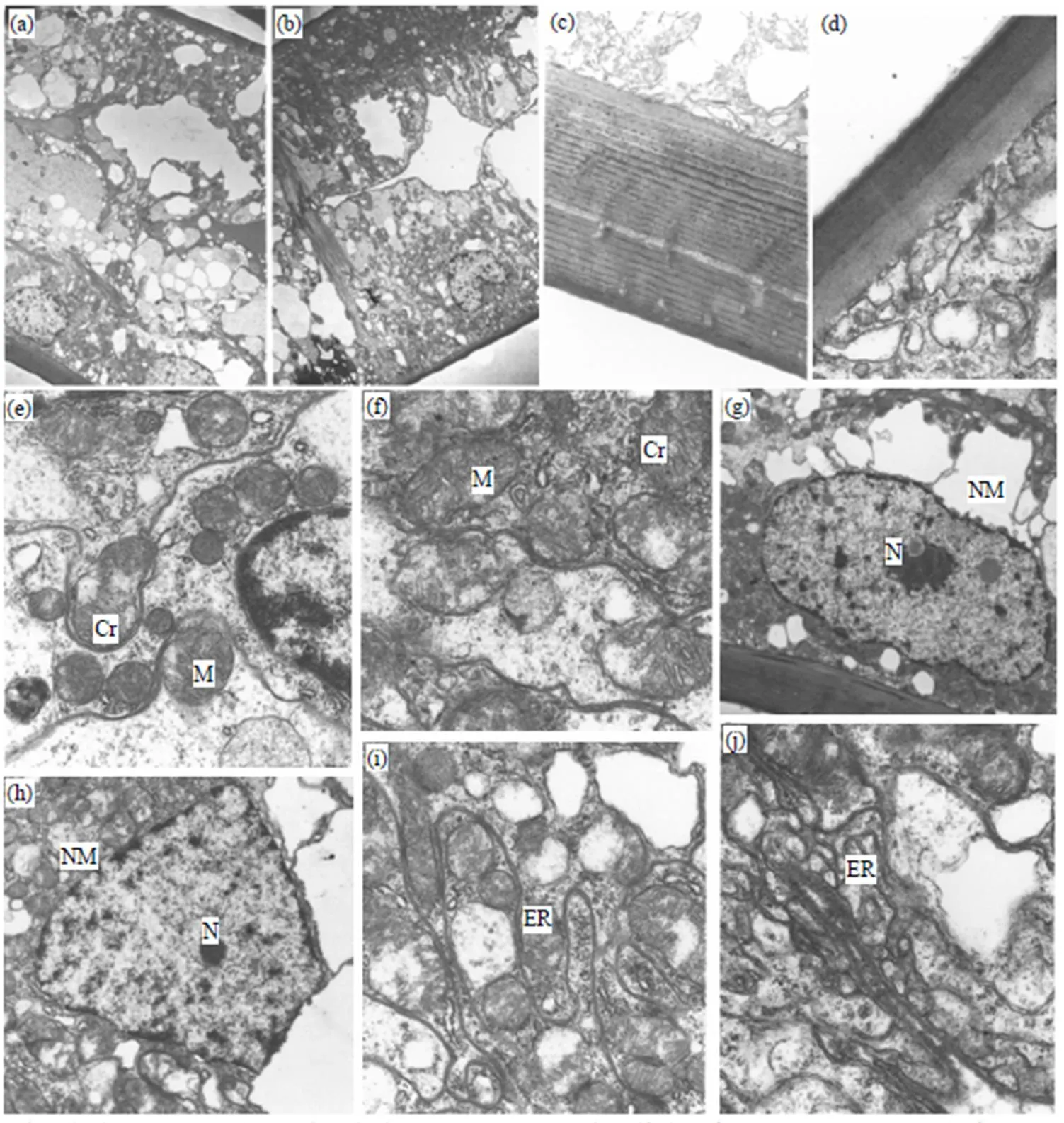
(a) 对照组鳃丝细胞(×2000); (b) 饥饿组鳃丝细胞(×2000); (c) 对照组鳃丝管壁角质层(×8000); (d) 饥饿组角质层(×25000); (e) 对照组线粒体及内嵴(×15000); (f) 饥饿组线粒体(×20000); (g) 对照组细胞核及核膜(×4000); (h) 饥饿组细胞核及核膜(×6000); (i) 对照组内质网(×15000); (j) 饥饿组内质网(×20000). M: 线粒体; Cr: 线粒体内嵴; NM: 细胞核核膜; N: 细胞核; ER: 内质网.
如图2所示, 对照组日本蟳肝胰腺肝管微绒毛丰富、排列致密; 线粒体靠近微绒毛区分布, 内嵴发达; 细胞质中粗面内质网和滑面内质网十分丰富, 滑面内质网呈环状排列; 细胞核规则, 结构完整, 核膜清晰; 脂肪滴电子密度均匀. 饥饿组日本蟳肝胰腺细胞器溶解得比较多, 空泡化比较严重. 部分肝管的微绒毛出现退化现象, 排列不致密, 不规则, 部分缺失; 线粒体比较丰富、完整, 结构清楚; 内质网数量仍然丰富, 但部分内质网出现水肿、扩张, 嵴有轻微破坏; 细胞核不规则, 染色质减少、空泡化; 脂肪滴边缘出现裂纹.

(a) 对照组肝管微绒毛(×8000); (b) 饥饿组肝管微绒毛(×10000); (c) 对照组线粒体(×10000); (d) 饥饿组线粒体(×10000); (e)对照组内质网(×3000); (f) 饥饿组内质网(×3000); (g) 对照组细胞核及核膜(×6000); (h) 饥饿组细胞核及核膜(×4000); (i) 对照组脂肪滴(×2500); (j) 饥饿组脂肪滴(×3000). Mv: 肝管微绒毛; M: 线粒体; ER: 内质网; NM: 细胞核核膜; N: 细胞核; Li: 脂肪滴.
如图3所示, 对照组日本蟳心脏心肌细胞排列规则、致密, 明暗带清晰; 线粒体和内质网丰富, 结构完整; 细胞核规则, 核膜清晰、完整, 异染色质丰富, 染色较深; 核糖体丰富. 16d饥饿胁迫后日本蟳心脏细胞超微结构出现轻微的变化. 肌原纤维排列有些疏松, 明暗带模糊; 线粒体部分解体, 空泡增多, 线粒体内嵴肿胀, 排列不规则, 有溶解现象; 内质网扩张, 空泡增多, 其他细胞器基本无变化.

(a) 对照组中的肌原纤维(×8000); (b) 饥饿组肌原纤维(×15000); (c) 对照组线粒体及内嵴(×12000); (d) 饥饿组线粒体(×12000); (e) 对照组细胞核及核膜(×8000); (f) 饥饿组细胞核及核膜(×8000); (g) 对照组核糖体(×6000); (h) 饥饿组核糖体(×6000). My: 肌原纤维; M: 线粒体; Cr: 线粒体内嵴; NM: 细胞核核膜; N: 细胞核; R: 核糖体.
如图4所示, 对照组中肌原纤维致密, 排列规则, 明暗带清晰; 线粒体丰富, 为椭圆形, 内嵴发达, 分布均匀, 间质浓密; 细胞核规则, 核膜完整. 肌肉细胞在16d饥饿胁迫后除了细胞核染色质固缩外, 其他细胞器无明显的变化.
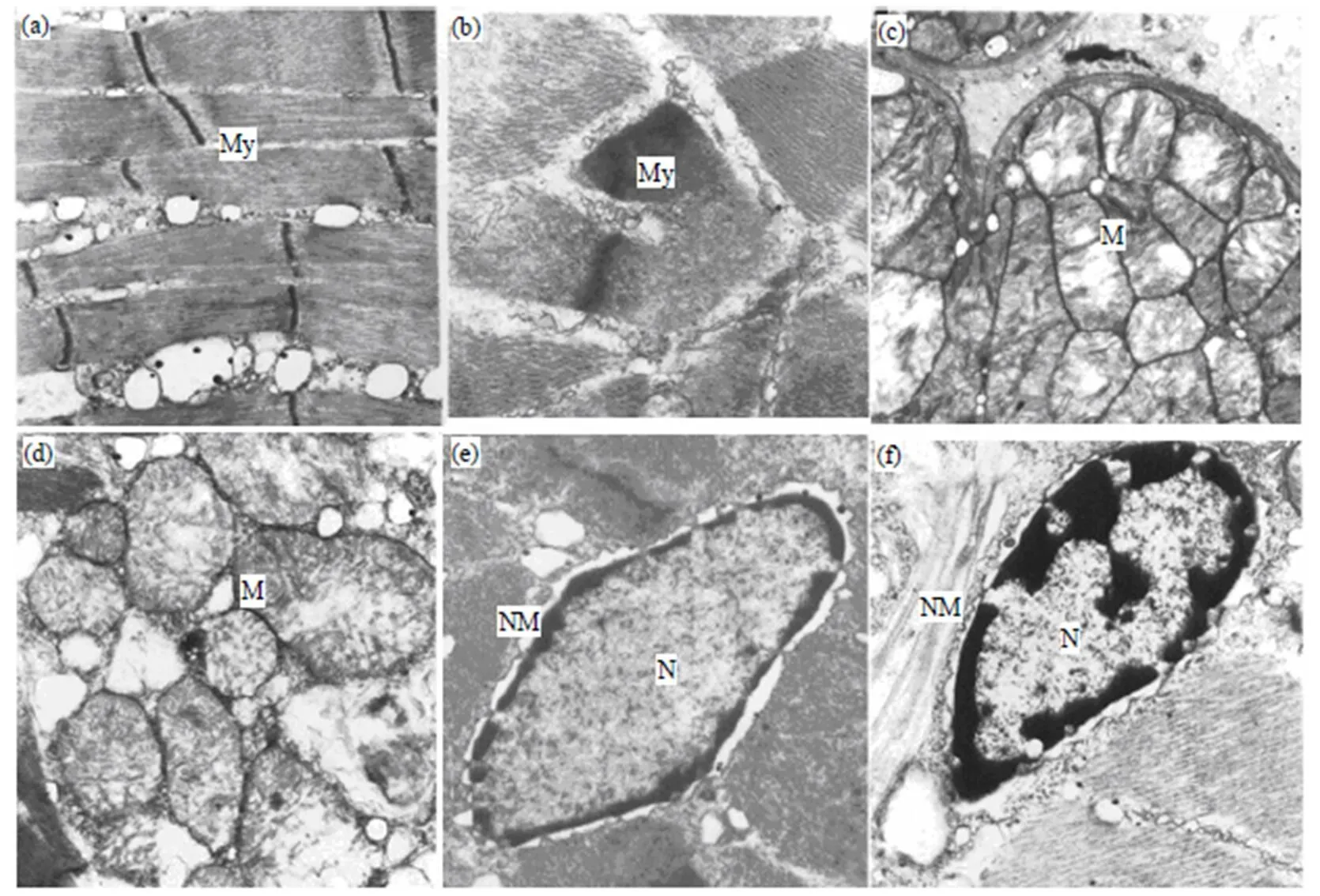
(a) 对照组肌原纤维(×10000); (b) 饥饿组肌原纤维(×10000); (c) 对照组线粒体(×7500); (d) 饥饿组线粒体(×12000); (e) 对照组细胞核及核膜(×15000); (f) 饥饿组细胞核及核膜(×8000). My: 肌原纤维; M: 线粒体; NM: 细胞核核膜; N: 细胞核.
3.1 饥饿胁迫下日本抗氧化酶的反应
学者前期研究表明抗氧化酶与生物机体的健康状况、免疫保护、环境应激等因素有关, 与生物抵抗环境胁迫密切相关, 既可以用来指示生物机体的非特异性免疫能力, 也可以用来作为环境胁迫检测的重要指标之一[10-11]. 水生生物在受到饥饿或其他环境因素胁迫时, 大量活性氧就会在细胞内累积, 活性氧在对抗外界胁迫的同时也会对细胞造成一定的伤害. 包括SOD、CAT在内的抗氧化酶体系可以降低自由基反应. SOD可以催化生物体内超氧阴离子(O2ˉ), 使之发生歧化反应生成过氧化氢(H2O2)和分子氧(O2), CAT能还原H2O2转化为水和氧, 两者联合作用在动态平衡中抵御自由基的损伤[12-14]. 由此可见, SOD和CAT在应对环境胁迫时起主导作用, 通过酶活性来调节代谢水平、能量分配和能源物质消耗以维持机体的正常生理活动[8].
本试验也证明饥饿胁迫下日本蟳4种组织中SOD和CAT参与了机体的生理调节, 而且不同组织中两种酶反应规律有所不同, 体现出细胞受到饥饿胁迫的破坏程度不同. 鳃的反应最快, 第4d两种酶就出现了较大的刺激和抑制效应, 肝胰腺中SOD在第12d才表现出强烈的刺激效应. 试验结束时鳃和肝胰腺中两种酶都没有恢复到对照组水平, 说明是饥饿胁迫的重要靶器官. 原因可能是鳃具有呼吸、排泄、调节渗透压等多种功能, 肝胰腺则是日本蟳营养消化吸收的场所, 对营养和能量的需求非常高, 所以对饥饿胁迫比较敏感[15]. 而心脏和肌肉不具有吸收和代谢功能, 所以两种酶变化程度不大.
此外, 同一组织中SOD和CAT活性受胁迫时的反应并不同步, 或许存在互补作用, 这和许星鸿等[16]的研究结果一致. 一方面是因为SOD和CAT是一个动态的保护系统, 两种酶互相配合, 共同抵御外界的侵害. 例如肝胰腺CAT活力降低的时候, SOD活力却显著升高. 另外一方面也可能是因为不同组织承担的生理功能不同, 饥饿胁迫所攻击的靶细胞不同.
3.2 饥饿胁迫对日本细胞超微结构的影响
细胞结构是动物生理功能的单位, 结构的损伤必将导致功能的变化, 功能异常又可能加速结构的损伤. 试验表明16d饥饿胁迫下日本蟳鳃细胞、肝胰腺细胞和心脏细胞的细胞器超微结构有不同的变化; 肌肉细胞和对照组比较除了细胞核染色质固缩外基本无变化; 心脏细胞中线粒体和肌原纤维是胁迫的主要靶部位, 其他细胞器基本无变化.

日本蟳具有一定的抵御饥饿胁迫的能力, 但这种能力是有限的. 组织细胞酶活性的大小是环境胁迫作用和保护酶系统自身防御共同作用的结果[22]. 在饥饿胁迫下的日本蟳可以通过调节抗氧化酶的水平来增强其清除活性氧的能力, 从而减轻外界胁迫的伤害, 但胁迫加强会导致日本蟳细胞代谢失调和细胞结构的破坏. 传统水产养殖主要依靠抗生素和化学药物进行病害防治, 不仅会污染水环境, 还会通过食物链影响人类健康. 可以考虑通过短期饥饿胁迫以及在饵料中添加小球藻等方法来激发日本蟳的抗氧化能力, 减少病虫害的大规模发生[23-24].
[1] 吴常文, 王志铮, 王伟洪, 等. 舟山近海日本蟳生物学、资源分布以及开发利用[J]. 浙江水产学院学报, 1998(1):13-18.
[2] 俞存根, 宋海棠, 姚光展. 东海日本蟳的数量分布和生物学特性[J]. 上海水产大学学报, 2005, 14(1):40-45.
[3] Hossain A M, Datta H M. Food deprivation induces differential changes in contents and microstructures of digestive tract and appendages in bluegill fish,[J]. Comparative Biochemistry and physiologypart A: physiology, 1991, 100(3):769-772.
[5] 卢俊姣, 刘淑兰, 翟少伟. 饥饿胁迫对罗非鱼肝胰脏抗氧化能力的影响[J]. 中国农学通报, 2013, 29(35):75-79.
[6] 金鑫, 徐钢春, 杜富宽, 等. 饥饿胁迫对刀鲚形体、体成分及血液生化指标的影响[J]. 动物学杂志, 2014, 49(6):897-903.
[7] 王东东, 吴成宾, 郑国栋, 等. 饥饿胁迫对团头鲂鳃组织结构及Na+/K+-ATP酶、抗氧化酶的影响[J]. 上海海洋大学学报, 2019, 28(5):765-771.
[8] 赵旺, 温为庚, 谭春明, 等. 饥饿胁迫对猛虾蛄不同组织免疫酶活性的影响[J]. 中山大学学报(自然科学版), 2021, 60(4):26-33.
[9] Beauchamp C, Fridovich I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels[J]. Analytical Biochemistry, 1971, 44(1):276-287.
[10] 江水恒, 王丹丽, 邹秀, 等. 不同阶段三疣梭子蟹保护酶及消化酶活性研究[J]. 河北渔业, 2012(4):9-12.
[11] Zhang H X, Pan L Q, Miao J J, et al. Effects of mercuric chloride on antioxidant system and DNA integrity of the crab[J]. Journal of Ocean University of China, 2009, 8(4):416-424.
[12] 丁金强, 刘萍, 李健, 等. 不同地理群体日本蟳非特异性免疫及抗氧化酶活力的比较[J]. 水产学报, 2013, 37(2):275-280.
[13] Legras S, Mouneyrac C, Amiard J C, et al. Changes in metallothionein concentrations in response to variation in natural factors (salinity, sex, weight) and metal contamination in crabs from a metal-rich estuary[J]. Journal of Experimental Marine Biology and Ecology, 2000, 246(2):259-279.
[14] Mouneyrac C, Amiard-Triquet C, Amiard J C, et al. Comparison of metallothionein concentrations and tissue distribution of trace metals in crabs () from a metal-rich estuary, in and out of the reproductive season[J]. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 2001, 129(3):193-209.
[15] 郑志仁, 王炳森, 蒋学之, 等. 环境病理学[M]. 济南: 山东科学技术出版社, 1991.
[16] 许星鸿, 张雁秋, 阎斌伦, 等. 氨氮胁迫对日本蟳免疫生理指标及器官结构的影响[J]. 生态学报, 2014, 34(14):3885-3894.
[17] 王永生. 饥饿(限食)和重喂对三种不同适盐性鱼类形态形状和肝细胞超微结构的影响[D]. 汕头: 汕头大学, 2002.
[18] 殷帅文, 林学群, 陈洁辉. 饥饿和再恢复投喂对鲻鱼肝细胞超微结构的影响[J]. 井冈山大学学报(自然科学版), 2011, 32(3):60-64.
[19] 张升利, 梁拥军, 孙向军, 等. 饥饿对星斑川鲽消化器官形态结构和组织学的影响[J]. 中国水产科学, 2012, 19(3):445-452.
[20] Weis P. Hepatic ultrastructure in two species of normal, fasted and gravid teleost fishes[J]. The American Journal of Anatomy, 1972, 133(3):317-331.
[22] 肖泽恒, 甘甜, 秦钟, 等. 饥饿胁迫对福寿螺生长、抗氧化系统及生化物质的影响[J]. 中国生态农业学报(中英文), 2022, 30(6):1036-1044.
[23] 王洪斌, 金学萍, 肖龙海, 等. 富硒海洋小球藻对三疣梭子蟹血清中部分免疫活性酶的影响[J]. 湖北农业科学, 2016, 55(14):3687-3689.
[24] 王家新, 苏宏, 王卫兵, 等. 海水小球藻粗多糖对日本蟳免疫相关酶活性的影响[J]. 水产科学, 2021, 40(6): 917-921.
Effects of starvation stress on the activities of antioxidant enzymes and cell ultrastructure of
DING Aixia
( School of Civil and Environmental Engineering, Ningbo University, Ningbo 315211, China )
Using the transmission electron microscopy and spectrophotometry, the current study was designed to investigate the effect of the starvation stress on the activities of superoxide dismutase (SOD), catalase (CAT) and the ultrastructure of 4 tissues (gill, hepatopancreas, heart and muscle) in. The results indicated that SOD and CAT activities inexhibited high tissue specificity. The order of SOD activity distributed in different tissues from high to low was heart, gill, muscle and hepatopancreas. The order of CAT activity was hepatopancreas, heart, gill and muscle. SOD and CAT activities have been changed , following the introduction of the starvation stress. SOD holistically showed a significant stimulating effect within 16 days. CAT in gill was alleviated and CAT in hepatopancreas showed initial decrease and subsequent increase. The ultrastructure of four tissue cells were significantly altered under the starvation stress. Swelling gill filament, thinning cuticle and partially dissolved mitochondria was evident in Gill cells. Decreased microvilli, mitochondrion partly disintegrated, expanded endoplasmic reticulum and a few irregular nuclei was found in hepatopancreas cells. Mitochondria in heart cells was partially dissolved and myofibrils were swelled. Chromatin of muscle cells were pagans. It is concluded thatitself has limited defense ability against starvation stress.
; starvation; antioxidant enzyme; ultrastructure
2022−03−04.
宁波大学学报(理工版)网址: http://journallg.nbu.edu.cn/
丁爱侠(1974-), 女, 江苏连云港人, 高级实验师, 主要研究方向: 海洋环境生态. Email: dingaixia@nbu.edu.cn
S917
A
1001-5132(2022)04-0001-08
(责任编辑 韩 超)
猜你喜欢 细胞核线粒体饥饿 不同组织来源线粒体提取效率和质量的差异研究重庆理工大学学报(自然科学)(2022年9期)2022-10-26线粒体自噬在纤维化疾病中作用的研究进展中华实用诊断与治疗杂志(2022年1期)2022-08-31指向科学思维的“细胞核的结构与功能”教学设计中学生物学(2022年3期)2022-05-13人参bZIP基因家族生物信息学分析中草药(2022年9期)2022-05-06向着“零饥饿”的目标金桥(2021年10期)2021-11-05MFN2调控线粒体动态变化的研究进展中国医药导报(2019年13期)2019-06-20“细胞核—系统的控制中心”一节的教学设计新教育时代·教师版(2018年48期)2018-01-24饥饿奏鸣曲语文世界(初中版)(2017年6期)2017-06-15为什么我们还保留着线粒体DNA?飞碟探索(2016年4期)2016-04-07饥饿的歌声小说月刊(2014年8期)2014-04-19







